

.
Bands result from membrane strip incubation.")
.
Bands result from membrane strip incubation.")
 and mouse anti-beta tubulin Ab(T0023) for 1 hour at 37°C. An AlexaFluor594 conjugated goat anti-rabbit IgG(H+L) Ab(Red) and an AlexaFluor488 conjugated goat anti-mouse IgG(H+L) Ab(Green) were used as the secondary antibody.
The nuclear counter stain is DAPI(blue).")
 and mouse anti-beta tubulin Ab(T0023) for 1 hour at 37°C. An AlexaFluor594 conjugated goat anti-rabbit IgG(H+L) Ab(Red) and an AlexaFluor488 conjugated goat anti-mouse IgG(H+L) Ab(Green) were used as the secondary antibody.
The nuclear counter stain is DAPI(blue).")

 microscopy at x200 magnification was used to assess cell morphology. The A549 cells (parental cells) had an epithelioid, rounded cobblestone appearance and there was limited formation of pseudopodia. A549/PTX and A549/DDP cells exhibited a spindle-shaped morphology and an increased formation of pseudopodia, indicating a loss of cell polarity. (B) E-cadherin, β-catenin, vimentin, MMP-2 and MMP-9 which are EMT-related proteins, were assessed in terms of expression levels. EMT-related transcription factors (Snail, Slug, Twist and ZEB1) were measured in A549/PTX and A549/DDP cells using western blot analysis. (C) The expression changes were confirmed at the mRNA level by qRT-PCR. Expression was standardized to the expression of GAPDH and normalized to 1.0 in the parental cells (compared with the parental A549 cells, means ± SEM, n=3, * P<0.05)")
 markers and b-catenin was detected at each time point. (A) Immunoblotting analysis of human primary cultured endometrial epithelial cell extracts using the corresponding antibodies. The ratios of each protein relative to non-treated cells were normalized to GAPDH. (B) The relative expression of HIF-1a, N-cadherin, E-cadherin, b-catenin, vimentin and snail proteins in human endometrial epithelial glands under hypoxic conditions at each time point was investigated by western blot. Data are represented as mean+SD and are representative of the relative expression of protein normalized by GAPDH. All experiments were repeated four times. Data were evaluated by one-way ANOVA analysis (*P , 0.05, **P , 0.01 compared with untreated group). (C) The changed cellular morphologies of human endometrial epithelial glands in hypoxia compared with cells in normoxia, the hypoxic time was 48 h. Red arrows indicate the spindle-shaped and fibroblast-like cells.")
, b-catenin (D–F), E-cadherin (G–I), N-cadherin (J–L) and vimentin (M–O) in normal endometrium (A, D, G, J, M), eutopic endometrium (B, E, H, K, N) and ovarian endometriosis (C, F, I, L,O). (P)Colon cancer tissue that was positive for HIF-1a. (Q) Healthy liver tissue that was negative for HIF-1a. (R) Peptide-blocking reagent without antibody was applied as the negative controls. Photographs were taken at magnifications of ×200 (left panels) and ×400 (right panels). N, normal endometrium; U, eutopic endometrium; E, ovarian endometriosis.")
 The levels of cyclin A, cyclin B, cyclin D1, and CDK2 were reduced in A375 cells transfected with OVOS2-shRNA; (b) The downregulated expression of N-cadherin accompanied with the upregulated expression of E-cadherin and β-catenin were observed in A375 cells transfected with OVOS2-shRNA; (c) The expression of p-FAK, p-AKT, and p-ERK were reduced in A375 cells transfected with OVOS2-shRNA; (d) The increased production of MMP-2 was observed in A375 transfected with OVOS2-shRNA; (e) GAPDH was used as the reference.")
Immunofluorescence assay showed that POLR1D overexpression increased the total β-catenin and the nuclear β-catenin levels in RKO cells, while POLR1D knockdown resulted in the opposite effects in LOVO cells.")
 Western blot showing the expression of wingless/integrated 1 (Wnt1), Wnt3a, Wnt5a, β‐catenin, and glyceraldehyde‐3 phosphate dehydrogenase (GAPDH) in five groups (n = 3). G1: negative control group; G2: running group; G3: weight‐bearing group; G4: positive control group; Neonatal: the neonatal bone, control.")
Protein expression level of P120 and β-catenin 24 h after HI brain injury.")
 Quantitative analysis results and representative images of the western blot results for -catenin and C-myc in HepG2 cells.")



 Representative western blots and
quantification data for BDNF/β-Actin, PI3K/β-Actin, p-TrkB/TrkB, p-AKT/AKT and p-CREB/CREB in each group, columns represent the mean ± SD, n = 5. (I) Double
staining for Claudin-5, Occludin, p120-Catenin, and β-Catenin/CD31/Hoechst. Red: Claudin-5/Occludin/p120-Catenin/β-Catenin; green: CD31; blue: Hoechst. Scale
bar, 20 μm #p < 0.05 for the M group versus the S group, *p < 0.05 for the TM group versus the M group. (
#p, *p < 0.05; ##p, **p < 0.01; ###p, ***p < 0.001).")
 Representative western blots and quantification data of TJ and AJ proteins in M, IM groups.
Columns represent mean ± SD, n = 5. (C) Claudin-5, Occludin, p120-Catenin, β-Catenin/CD31/Hoechst staining of sections from the spinal cord in M, IM groups. Red:
Claudin-5/Occludin/p120-Catenin/β-Catenin; green: CD31; blue: Hoechst. Scale bar, 20 μm. (
$
p < 0.05; $$p < 0.01; $$$p < 0.001).")
; CMTM5 was highly expressed in adjacent tissues (p<0.05).")
 A498 cell invasion and (B) 769-P cell invasion. In the two panels, data are presented as the mean ± SD of the number of cells in five randomly selected fields of view from three independent experiments (magnification, ×200). (C) Effect of SEC61G knockdown on E-cadherin, N-cadherin and β-catenin protein expression levels in A498 cells. (D) Effect of SEC61G knockdown on E-cadherin, N-cadherin and β-catenin protein expression levels in 769-P cells. Data are presented as the mean ± SD of three independent experiments. **P<0.01 and ***P<0.001 vs. NC. SEC61G, SEC61 translocon subunit-γ; si, small interfering RNA; NC, negative control; E-cad, E-cadherin; N-cad, N-cadherin.")
 and ICG‐001 (5, 10 and 15 μM/ml) for 24, 48 and 72 h. The viability of cells was estimated using the CCK‐8 assay. C, IF assay for KI67 in J774A.1 cells. D, E, Western blotting analysis and their respective quantitation showing the protein expression of FKN, β‐catenin, Wnt‐4, c‐myc and cyclinD1 in J774A.1 cells. F, The secretion of cyclinD1 in J774A.1 cell supernatants was detected using ELISA. * P < .05 compared with the control group; # P < .05 compared with the LPS group. G, The subcellular localization of cyclin D1 was identified by immunostaining using anti‐Cyclin D1 and observed using confocal microscopy. Scale bars represent 10 μm")
 and SP-B through the Wnt signaling pathway. After transfection of mir-296-5p overexpression and empty vectors in the A549 cell line, the expression of WNT7B, β-catenin, SP-A, and SP-B was assessed using western blotting.")
 The protein levels of p-AKT,
AKT, p-GSK3β, GSK3β, p-β-catenin, and β-catenin in lung tissues of mice. (b) Immunohistochemistry staining of β-catenin. Red arrow: the
expression of β-catenin in nucleus. Scale bar
=50 μm. Data were expressed as mean ± SD. n
= 6 in each group. **p < 0.01 vs. control; #p <
0.05, ##p < 0.01 vs. BLM.")
. To fit into the manuscript properly, the gel was reasonably trimmed. aa, P < 0.01 vs the control group; b, P < 0.05, bb, P < 0.01 vs the Wnt-1 group. ##, P < 0.01 vs the normal group; *, P < 0.05 and **, P < 0.01 vs the model group")
 The extracellular acidification rate of the LAD cells transfected by si-ALDOA and si-AC122108.1. (E,F) GSK-3β interacts with ALDOA proteins by co-immunoprecipitation with anti-FLAG M2 beads. (G-J) The differential expression of β-catenin and p-β-catenin in LAD cells transfected by si-ALDOA and si-AC122108.1. (K-N) The differential expression of EMT (pithelial-mesenchymaltransition)-related proteins and downstream-target proteins of the WNT/β-Catenin signaling pathway in LAD cells transfected by si-ALDOA and si-AC122108.1. (O,P) The differential expression of β-catenin and p-β-catenin in LAD cells transfected by oe-ALDOA and si-AC122108.1. (Q,R) The differential expression of EMT-related proteins and downstream target proteins of the WNT/β-Catenin signaling pathway in LAD cells transfected by oe-ALDOA and si-AC122108.1 (*P<0.05, **P<0.01, ***P<0.001, ****P<0.0001). ALDOA, aldolase A; LAD, lung adenocarcinoma; EMT, epithelial-mesenchymal transition.")
 The expression of ALDOA and β-catenin in normal lung tissues and lung cancer tumor tissues with a low and high expression of AC122108.1 in LAD patients (scale bar: 50 µm; the white arrows indicate ALDOA nuclear translocation; the green arrows indicate β-catenin nuclear translocation). (B) The tissue immunofluorescence staining of ALDOA and β-catenin in normal brain tissues and brain cancer tumor tissues with a low and high expression of AC122108.1 in LAD patients (scale bar: 50 µm; the white arrows indicate ALDOA nuclear translocation; the green arrows indicate β-catenin nuclear translocation). ALDOA, aldolase A; LAD, lung adenocarcinoma.")
 Violin plot of gene expression; (B) Based on the results of differential analysis, we screened genes with FDR < 0.05 and |log2FC|>1 as significantly differential genes, and the statistical histogram of differential genes. (C) The different genes volcano plot of LPS + siR-A33 vs LPS; (D) KEGG enrichment: the top 20 pathways with the smallest Q value. Inflammation-related pathways are in the red box, and pathways in which Wnt signaling first appears are in the blue box; (E) Heatmap of gene expression associated with wnt/β-catenin signaling; (F) Western blot analysis of Axin2 and β-catenin protein levels after down-regulation of A33. ***P < 0.001 vs LPS group.")
 Immunohistochemistry results for Axin2 and β-catenin in each group; (B,C) Real-time PCR and WB results confirmed that mRNA and protein levels of Axin and β-catenin decreased significantly in HDG treatment group; (D) Immunohistochemistry results of Axin2 and β-catenin after knockdown of A33 in vivo; (E,F) Real-time PCR and WB results confirmed that HDG had a significantly inhibitory effect on Axin2 and β-catenin in LPS-induced pTEC; (G,H) Real-time PCR and WB confirmed that the expression of Axin2/β-catenin was synergistic with the expression of A33 after knockdown and over-expression in vitro. *P < 0.05, **P < 0.01, ***P < 0.001 vs Cis/LPS group; #P < 0.05, ###P < 0.001 vs Ctrl group.")
 The effects between circ_0051079 knockdown and TRIM66 overexpression on the production of β-catenin, c-myc and cyclinD1 were determined by Western blot analysis. *P < 0.05.")

. The protein-protein interaction (PPI) network of the KRT17 from STRING online database (https://string-db.org) was constructed (b). The protein levels in AMC-HN-8 cells were evaluated using western blot (c). The protein levels in TU177 cells were estimated using western blot. ∗A significant difference compared with the sh-NC group. ∗∗P < 0.01 and ∗∗∗P < 0.001.")
 HMGA2 protein expression significantly decreased following transfection with siRNA-HMGA2. (B) HMGA2 mRNA expression significantly decreased following transfection with siRNA-HMGA2. (C) Wnt protein expression significantly decreased following transfection with siRNA-Wnt. (D) Wnt mRNA expression significantly decreased following transfection with siRNA-Wnt. (E) β-catenin protein expression significantly decreased following transfection with siRNA-β-catenin. (F) β-catenin mRNA expression significantly decreased following transfection with siRNA-β-catenin. *P")

 can activate the wnt/β-catenin signaling pathway. A: functional enrichment analysis of the results of protein spectrum identification. B: the results of KEGG analysis showed that the signal pathway related to the DEPDC1B interacting proteins. C and D: DEPDC1B is involved in the nuclear translocation of β-catenin. Hs578T cells (C) were transfected with negative control (NC) and DEPDC1B siRNA, MDA-MB-157 cells (D) were transfected with vector and DEPDC1B overexpression plasmids, and the expression of β-catenin was detected by immunofluorescence after 48 h. Scale bars, 20 μm. E: the results of the statistical analysis of C and D. Analysis between 2 groups was conducted by unpaired Student’s t test. *P < 0.05. F: the expression levels of wnt3a, phospho (p-)GSK-3β (ser9), GSK-3β, and β-catenin were detected by Western blot, and GAPDH was used as the internal control. G: Western blot results of the expression of DEPDC1B, wnt3a, and β-catenin in MDA-MB-157 cells transfected with DEPDC1B overexpression plasmid alone or in combination with XAV-939, with GAPDH as the internal control.")
 mediates the deubiquitination of β-catenin by ubiquitin-specific protease 5 (USP5). A: coimmunoprecipitation (Co-IP) results show that DEPDC1B interacts with β-catenin in breast cancer cells. MDA-MB-157 cells were transfected with FLAG-DEPDC1B and HA-β-catenin alone or in combination, and Co-IP was used to detect DEPDC1B and β-catenin. B: Co-IP results show that DEPDC1B interacts with USP5 in breast cancer cells. MDA-MB-157 cells were cotransfected with FLAG-DEPDC1B and MYC-USP5 plasmids, and DEPDC1B and USP5 were detected by Co-IP. C: DEPDC1B, USP5, and β-catenin were colocalized in breast cancer cells by detection through transfection of DEPDC1B, USP5, and β-catenin in MDA-MB-157 cells. Scale bars, 60 μm. D: overexpression of DEPDC1B weakens the effect of USP5 on β-catenin protein degradation level. After MDA-MB-157 cells were transfected with USP5 alone or in combination with the DEPDC1B overexpression plasmid and treated with cycloheximide (CHX) (0, 4, 8, 12 h), β-catenin was detected by Western blotting, and β-actin was used as an internal control. NC, negative control. E: knockdown of DEPDC1B promotes the ubiquitination of β-catenin, and combining overexpression of USP5 decreases the ubiquitination of β-catenin in breast cancer cells. MDA-MB-157 cells were treated with DEPDC1B siRNA combined with MYC-USP5, HA-β-catenin, and His-Ub and then treated with MG132 to prepare cell extracts. Anti-His was used for immunoblot (IB) analysis.")
 C4-2 cells were implanted subcutaneously in male BALB/c mice. When tumors became palpable, mice administered daily by oral gavage either with vehicle (10% DMSO in PBS) or NCB0846 (80 mg/kg of body weight) for 10 days (n = 4 mice for each treatment). Tumor volumes were measured with calipers. (B) Tumor size of xenografts of the above represented the growth of tumor over 10 days (n = 4) in athymic nude mice (p < 0.001). Data are shown as mean ± SD. (C) Tumor weight of the control mice tumors and NCB-0846-treated mice tumors (p < 0.001). Data are shown as mean ± SD. (D) Body weight of nude mice after implantation of control or C4-2 xenografts and treatment with vehicle or NCB-0846 for 4 weeks. (E) Quantitation of Ki-67, TNIK, p-EGFR, β-catenin, vimentin, E-cadherin, BMP6, and BMP7 expressions in C4-2 xenograft tumors from each group; specimens were got at 10 days posttreatment. Scale bars: 500 μm. The IHC was scored according to number of cells expressing the indicated proteins, and statistical analysis was performed (non-parametric Kruskal-Wallis test) in order to determine significance. Data are shown as mean ± SD.")
 was positively expressed, while keratin (red) was negatively expressed. The morphology of HConFs in bright-field presents as the typical spindle shape (right panel). B: Volcano plots of differentially expressed proteins (DEPs) between the TGFβ1 and control (CON) groups. C: KEGG enrichment analysis of DEPs between the TGFβ1 and CON groups. Significant enrichment of the KEGG pathways are visualized by bubble maps. The boxed region indicates the has04310 Wnt-signaling pathway. D: Protein intensities of LRP5/6 and CTNNB1, as detected by liquid chromatography–tandem mass spectrometry. Levels of LRP5/6 and CTNNB1 are significantly improved in TGFβ1-induced HConFs E: mRNA levels of members of the Wnt pathway (WNT3A, LEF1, CTNNB1, AXIN2, SFRP1, LRP5, and LRP6). F and G: Representative images (F) and quantitative analysis (G) of fluorescence-labeled β-catenin. TGFβ1 promotes the nuclear translocation of β-catenin. The arrows indicate nuclear-located β-catenin. H and I: Nuclear distribution of β-catenin, as detected by Western blot analysis. TGFβ1 increases the expression of β-catenin in the nucleus. Data are expressed as means ± SD. n = 3 (D and E); n = 5 (I); n = 8 (G). ∗P < 0.05, ∗∗P < 0.01, and ∗∗∗P < 0.001. Scale bars = 50 μm. Original magnification, ×100 (A, bright-field image). AXIN, axis inhibition protein; CTNN, catenin; ECM, extracellular matrix; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; KEGG, Kyoto Encyclopedia of Genes and Genomes pathways; LEF, lymphoid enhancer–binding factor; LRP, low-density lipoprotein receptor–related protein; SFRP, secreted frizzled-related protein.")
 was positively expressed, while keratin (red) was negatively expressed. The morphology of HConFs in bright-field presents as the typical spindle shape (right panel). B: Volcano plots of differentially expressed proteins (DEPs) between the TGFβ1 and control (CON) groups. C: KEGG enrichment analysis of DEPs between the TGFβ1 and CON groups. Significant enrichment of the KEGG pathways are visualized by bubble maps. The boxed region indicates the has04310 Wnt-signaling pathway. D: Protein intensities of LRP5/6 and CTNNB1, as detected by liquid chromatography–tandem mass spectrometry. Levels of LRP5/6 and CTNNB1 are significantly improved in TGFβ1-induced HConFs E: mRNA levels of members of the Wnt pathway (WNT3A, LEF1, CTNNB1, AXIN2, SFRP1, LRP5, and LRP6). F and G: Representative images (F) and quantitative analysis (G) of fluorescence-labeled β-catenin. TGFβ1 promotes the nuclear translocation of β-catenin. The arrows indicate nuclear-located β-catenin. H and I: Nuclear distribution of β-catenin, as detected by Western blot analysis. TGFβ1 increases the expression of β-catenin in the nucleus. Data are expressed as means ± SD. n = 3 (D and E); n = 5 (I); n = 8 (G). ∗P < 0.05, ∗∗P < 0.01, and ∗∗∗P < 0.001. Scale bars = 50 μm. Original magnification, ×100 (A, bright-field image). AXIN, axis inhibition protein; CTNN, catenin; ECM, extracellular matrix; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; KEGG, Kyoto Encyclopedia of Genes and Genomes pathways; LEF, lymphoid enhancer–binding factor; LRP, low-density lipoprotein receptor–related protein; SFRP, secreted frizzled-related protein.")

 Relative mRNA expression of Wnt pathway related genes was measured in dermal fibroblast cells 24 h after transfection with NC or miR-140-y mimics by qPCR. (B–C) Relative protein expression of Wnt pathway related genes was measured in dermal fibroblast cells 72 h after transfection with NC or miR-140-y mimics by Western blot. (D–E) Representative observations of the EdU assay of dermal fibroblast cells after transfection with NC and miR-140-y mimics (Bars: 100 µm). Red illustrate the EdU staining and blue display the cell nuclei stained with Hoechst 33342. (F) The mRNA expression of proliferation related gene (PCNA). (G-H) Protein expression level of proliferation related gene (PCNA). The data were shown as mean ± SEM, n = 3, * p < 0.05, ** p < 0.01.")
 Immunofluorescence staining of TFEB (GFP) and β-catenin (red) of the CFs co-incubated with TGF- β1 (5 ng/mL) for 12 h. The interaction between TFEB and β-catenin was showed using high resolution confocal microscopy analysis s. PNU-74654 (100 ng/mL) was used to for 12 h to inhibit β-catenin-TCF/LEF1 complex (40X). Scale bars represent 100 µm. (B) Immunofluorescence staining of TFEB (GFP) and β-catenin (red) of the CFs co-incubated with TGF- β1 (5 ng/mL) for 12 h. Si-TFEB was used to inhibit TFEB (40X). Scale bars represent 100 µm. (C) The expression of α-SMA was detected using western blotting. PNU-74654 (100 ng/mL) was used to for 12 h to inhibit β-catenin-TCF/LEF1 complex. (D) Cell viability assessed by CCK-8 assay after rat CFs were treated with TGF- β1 (5 ng/mL) for 12 h. PNU-74654 (100 ng/mL) was used to for 12 h to inhibit β-catenin-TCF/LEF1 complex. (E) Cell migration was evaluated by the Transwell assay for the CFs co-incubated with TGF- β1 (5 ng/mL) for 12 h. PNU-74654 (100 ng/mL) was used to for 12 h to inhibit β-catenin-TCF/LEF1 complex. (F) Cell migration was evaluated by wound healing assay for the CFs co-incubated with TGF- β1 (5 ng/mL) for 24 h. PNU-74654 (100 ng/mL) was used to for 12 h to inhibit β-catenin-TCF/LEF1 complex (*: p < .05, **: p ¡ 0.01, ***: p < 0.001, ****: p < 0.0001). Scale bars represent 500 µm.")
 Cellular β-catenin localization in the SKOV3 cells was visualized using immunofluorescence staining (white circle indicated β-catenin nuclear localization). Scale bar = 50 µm. (b) Representative protein bands of β-catenin in cytoplasm from SKOV3 cells. GAPDH served as the internal reference. (c) Representative protein bands of β-catenin in nuclear from SKOV3 cells. Histone H3 served as the internal reference. Nor, normoxia; Hyp, hypoxia; shNC, negative control short hairpin RNA; sh1, ELK3 shRNA 1; sh2, ELK3 shRNA 2.")
 CTC-TJH-01 cells were treated with TSZAF mc (0, 0.5, 1 μmol·L−1) for 24 h,then the relative mRNA expression levels of E-cadherin, β-catenin, APC, Wnt-3α,c-Myc, Cyclin D1 and MMP-7 were analyzed by RT-PCR. (B) The CTC-TJH-01 and (C) LLC cells were exposed to TSZAF mc (0, 0.5, 1 μmol·L−1) for 24 h, the expression of E-cadherin, β-catenin, p-β-catenin, Wnt-3α, c-Myc, Cyclin D1 and MMP-7 was detected by WB. β-Actin was used as an internal standard. (D) The immunohistochemistry staining results of β-catenin expression in lung metastases. *P < 0.05, **P < 0.01, ***P < 0.001 vs the control group.")
 The optimal drug concentrations of RSL3 and Ferrostatin-1 were determined using CCK8 assays. (C,D) Relative protein expression levels of SLC7A11 and GPX4 under chronic hypoxia and after treatment with RSL3 and Ferrostatin-1. The original WB images are shown in Figure S9. (E) Immunofluorescence staining for the epithelial marker β-catenin and the fibroblast marker Vimentin, with DAPI staining for nuclei. Magnification ×200, scale bar = 100 μm. (F,G) Fluorescence intensity analysis of β-catenin and Vimentin under chronic hypoxia and after treatment with RSL3 and Ferrostatin-1. (H) Western blot bands for E-cadherin, α-SMA, Collagen I, and Fibronectin. The original WB images are shown in Figure S10. (I) Relative protein expression levels of E-cadherin, α-SMA, Collagen I, and Fibronectin in the different treatment groups. All data are presented as the mean ± SEM, ns = not significant, * p < 0.05; ** p < 0.01; *** p < 0.001.")

 Vero and IPEC-J2 cells were infected with GDgh strain (MOI=0.1) for 12, 24, 36 h, and (MOI=1.0) for 48, 60 h, respectively. The expression of β-catenin, MEF2A and PEDV N protein was detected by IB. (C) Vero cells were transfected with scrambled and a specific siRNA (monkey MEF2A) at a concentration of 50 nM, with or without infection with PEDV GDgh strain (MOI=0.01) for 24 h. The expression of GLUT1, GLUT3, β-catenin, MEF2A and PEDV N protein was detected by IB. (D and E) Vero cells were treated as A and B. The abundance of GLUT1 and GLUT3 mRNA was assessed by RT-qPCR, respectively, * *P")
 treatment. C Cell proliferation detected using CCK-8 assay. D Cell apoptosis detected using flow cytometric analysis. Adipogenic (E) and osteogenic (F) differentiation abilities of radiation-treated hucMSCs after BYSL upregulation. Scale: 50 μm. *p < 0.05 compared to control group; #p < 0.05, ###p < 0.001 compared to 1 Gy + oe-NC group; $p < 0.05, $$p < 0.01, $$$p < 0.001 compared to 1 Gy + oe-BYSL group")
 Principal Component Analysis (PCA) was performed on transcriptomic data to evaluate gene expression variations across DMSO controls, LPS-treated, and LPS + NorCA groups. (b) Volcano plot demonstrating NorCA-modulated gene expression changes in LPS-stimulated conditions. (c) Heatmap showing the expression levels of the top 20 upregulated and downregulated genes in the LPS group and LPS + NorCA group in the sequencing data. Gene Ontology (GO) enrichment analysis of Wnt pathway-associated DEGs in (d) biological processes, (e) cellular components, and (f) molecular functions. (g) Box plot of Ctnnb1 (Wnt/ β -catenin pathway) expression comparing LPS-treated and LPS + NorCA groups. (h) The expression levels of CTNNB1 in the LPS group and the LPS + NorCA group were detected by qPCR-RT. (i,j) Western blot analysis quantified β -catenin expression in LPS-treated versus LPS + NorCA groups.")
 in the Control group, marked increase in expression (black arrows) in L-MTZ, increased expression (black arrows) in S-MTZ. BMP-2: expression findings between the groups, moderate expression (black arrow) in the Control group, increase in expression (black arrows) in L-MTZ, marked expression (black arrow) in S-MTZ. Runx2: moderate expression (black arrows) in the Control group, increase in expression (black arrows) in L-MTZ, marked expression (black arrows) in S-MTZ. ALP: moderate expression (black arrow) in the Control group, marked increase in expression (black arrows) in L-MTZ, moderate expression (black arrow) in S-MTZ. OCN: moderate expression (black arrow) in the control group, markedly increased expression (black arrow) in L-MTZ, and increased expression (black arrow) in S-MTZ. RANKL: Similar RANKL expressions (black arrows) in (A) Control group, (B) L-MTZ, and (C) S-MTZ, Streptavidin biotin peroxidase method, Scale bars = 50 μm Representative histopathological images among the groups. (A) Marked fibrous tissue, moderate new bone formation (white arrow with a black border), and residual graft materials (RG) in the Control group. (B) Decreased fibrous tissue, moderate residual graft material (RG), and marked new bone formation (white arrow with a black border) in L-MTZ. (C) Decreased fibrous tissue, moderately increased new bone formation (white arrow with a black border), and residual graft materials (RG) in S-MTZ. HE, Scale bars = 200 μm")
 Western blot analysis of β-catenin, c-Myc, and SOX2 protein expression in NPC cells. (B) Spheroid formation assay to examine changes in stem-like properties.")
| Product: | beta Catenin Antibody |
| Catalog: | AF6266 |
| Description: | Rabbit polyclonal antibody to beta Catenin |
| Application: | WB IHC IF/ICC |
| Cited expt.: | WB, IHC, IF/ICC |
| Reactivity: | Human, Mouse, Rat |
| Prediction: | Pig, Zebrafish, Bovine, Horse, Sheep, Rabbit, Dog, Chicken, Xenopus |
| Mol.Wt.: | 80-100kD(Observed); 85kD(Calculated). |
| Uniprot: | P35222 |
| RRID: | AB_2835124 |
Control Products
Related Downloads
Protocols
Product Info
*The optimal dilutions should be determined by the end user. For optimal experimental results, antibody reuse is not recommended.
*Tips:
WB: For western blot detection of denatured protein samples. IHC: For immunohistochemical detection of paraffin sections (IHC-p) or frozen sections (IHC-f) of tissue samples. IF/ICC: For immunofluorescence detection of cell samples. ELISA(peptide): For ELISA detection of antigenic peptide.
Cite Format: Affinity Biosciences Cat# AF6266, RRID:AB_2835124.
Fold/Unfold
Beta catenin; Beta-catenin; Cadherin associated protein; Catenin (cadherin associated protein), beta 1, 88kDa; Catenin beta 1; Catenin beta-1; CATNB; CHBCAT; CTNB1_HUMAN; CTNNB; CTNNB1; DKFZp686D02253; FLJ25606; FLJ37923; OTTHUMP00000162082; OTTHUMP00000165222; OTTHUMP00000165223; OTTHUMP00000209288; OTTHUMP00000209289;
Immunogens
A synthesized peptide derived from human beta Catenin, corresponding to a region within N-terminal amino acids.
Expressed in several hair follicle cell types: basal and peripheral matrix cells, and cells of the outer and inner root sheaths. Expressed in colon. Present in cortical neurons (at protein level). Expressed in breast cancer tissues (at protein level) (PubMed:29367600).
- P35222 CTNB1_HUMAN:
- Protein BLAST With
- NCBI/
- ExPASy/
- Uniprot
MATQADLMELDMAMEPDRKAAVSHWQQQSYLDSGIHSGATTTAPSLSGKGNPEEEDVDTSQVLYEWEQGFSQSFTQEQVADIDGQYAMTRAQRVRAAMFPETLDEGMQIPSTQFDAAHPTNVQRLAEPSQMLKHAVVNLINYQDDAELATRAIPELTKLLNDEDQVVVNKAAVMVHQLSKKEASRHAIMRSPQMVSAIVRTMQNTNDVETARCTAGTLHNLSHHREGLLAIFKSGGIPALVKMLGSPVDSVLFYAITTLHNLLLHQEGAKMAVRLAGGLQKMVALLNKTNVKFLAITTDCLQILAYGNQESKLIILASGGPQALVNIMRTYTYEKLLWTTSRVLKVLSVCSSNKPAIVEAGGMQALGLHLTDPSQRLVQNCLWTLRNLSDAATKQEGMEGLLGTLVQLLGSDDINVVTCAAGILSNLTCNNYKNKMMVCQVGGIEALVRTVLRAGDREDITEPAICALRHLTSRHQEAEMAQNAVRLHYGLPVVVKLLHPPSHWPLIKATVGLIRNLALCPANHAPLREQGAIPRLVQLLVRAHQDTQRRTSMGGTQQQFVEGVRMEEIVEGCTGALHILARDVHNRIVIRGLNTIPLFVQLLYSPIENIQRVAAGVLCELAQDKEAAEAIEAEGATAPLTELLHSRNEGVATYAAAVLFRMSEDKPQDYKKRLSVELTSSLFRTEPMAWNETADLGLDIGAQGEPLGYRQDDPSYRSFHSGGYGQDALGMDPMMEHEMGGHHPGADYPVDGLPDLGHAQDLMDGLPPGDSNQLAWFDTDL
Predictions
Score>80(red) has high confidence and is suggested to be used for WB detection. *The prediction model is mainly based on the alignment of immunogen sequences, the results are for reference only, not as the basis of quality assurance.
High(score>80) Medium(80>score>50) Low(score<50) No confidence
Research Backgrounds
Key downstream component of the canonical Wnt signaling pathway. In the absence of Wnt, forms a complex with AXIN1, AXIN2, APC, CSNK1A1 and GSK3B that promotes phosphorylation on N-terminal Ser and Thr residues and ubiquitination of CTNNB1 via BTRC and its subsequent degradation by the proteasome. In the presence of Wnt ligand, CTNNB1 is not ubiquitinated and accumulates in the nucleus, where it acts as a coactivator for transcription factors of the TCF/LEF family, leading to activate Wnt responsive genes. Involved in the regulation of cell adhesion, as component of an E-cadherin:catenin adhesion complex. Acts as a negative regulator of centrosome cohesion. Involved in the CDK2/PTPN6/CTNNB1/CEACAM1 pathway of insulin internalization. Blocks anoikis of malignant kidney and intestinal epithelial cells and promotes their anchorage-independent growth by down-regulating DAPK2. Disrupts PML function and PML-NB formation by inhibiting RANBP2-mediated sumoylation of PML. Promotes neurogenesis by maintaining sympathetic neuroblasts within the cell cycle (By similarity).
Phosphorylation at Ser-552 by AMPK promotes stabilizion of the protein, enhancing TCF/LEF-mediated transcription (By similarity). Phosphorylation by GSK3B requires prior phosphorylation of Ser-45 by another kinase. Phosphorylation proceeds then from Thr-41 to Ser-37 and Ser-33. Phosphorylated by NEK2. EGF stimulates tyrosine phosphorylation. Phosphorylation on Tyr-654 decreases CDH1 binding and enhances TBP binding. Phosphorylated on Ser-33 and Ser-37 by HIPK2 and GSK3B, this phosphorylation triggers proteasomal degradation. Phosphorylation on Ser-191 and Ser-246 by CDK5. Phosphorylation by CDK2 regulates insulin internalization. Phosphorylation by PTK6 at Tyr-64, Tyr-142, Tyr-331 and/or Tyr-333 with the predominant site at Tyr-64 is not essential for inhibition of transcriptional activity.
Ubiquitinated by the SCF(BTRC) E3 ligase complex when phosphorylated by GSK3B, leading to its degradation. Ubiquitinated by a E3 ubiquitin ligase complex containing UBE2D1, SIAH1, CACYBP/SIP, SKP1, APC and TBL1X, leading to its subsequent proteasomal degradation (By similarity).
S-nitrosylation at Cys-619 within adherens junctions promotes VEGF-induced, NO-dependent endothelial cell permeability by disrupting interaction with E-cadherin, thus mediating disassembly adherens junctions.
O-glycosylation at Ser-23 decreases nuclear localization and transcriptional activity, and increases localization to the plasma membrane and interaction with E-cadherin CDH1.
Deacetylated at Lys-49 by SIRT1.
Cytoplasm. Nucleus. Cytoplasm>Cytoskeleton. Cell junction>Adherens junction. Cell junction. Cell membrane. Cytoplasm>Cytoskeleton>Microtubule organizing center>Centrosome. Cytoplasm>Cytoskeleton>Spindle pole. Cell junction>Synapse. Cytoplasm>Cytoskeleton>Cilium basal body.
Note: Colocalized with RAPGEF2 and TJP1 at cell-cell contacts (By similarity). Cytoplasmic when it is unstabilized (high level of phosphorylation) or bound to CDH1. Translocates to the nucleus when it is stabilized (low level of phosphorylation). Interaction with GLIS2 and MUC1 promotes nuclear translocation. Interaction with EMD inhibits nuclear localization. The majority of beta-catenin is localized to the cell membrane. In interphase, colocalizes with CROCC between CEP250 puncta at the proximal end of centrioles, and this localization is dependent on CROCC and CEP250. In mitosis, when NEK2 activity increases, it localizes to centrosomes at spindle poles independent of CROCC. Colocalizes with CDK5 in the cell-cell contacts and plasma membrane of undifferentiated and differentiated neuroblastoma cells. Interaction with FAM53B promotes translocation to the nucleus (PubMed:25183871).
Expressed in several hair follicle cell types: basal and peripheral matrix cells, and cells of the outer and inner root sheaths. Expressed in colon. Present in cortical neurons (at protein level). Expressed in breast cancer tissues (at protein level).
Belongs to the beta-catenin family.
Research Fields
· Cellular Processes > Cellular community - eukaryotes > Focal adhesion. (View pathway)
· Cellular Processes > Cellular community - eukaryotes > Adherens junction. (View pathway)
· Cellular Processes > Cellular community - eukaryotes > Signaling pathways regulating pluripotency of stem cells. (View pathway)
· Environmental Information Processing > Signal transduction > Rap1 signaling pathway. (View pathway)
· Environmental Information Processing > Signal transduction > Wnt signaling pathway. (View pathway)
· Environmental Information Processing > Signal transduction > Hippo signaling pathway. (View pathway)
· Human Diseases > Infectious diseases: Bacterial > Bacterial invasion of epithelial cells.
· Human Diseases > Infectious diseases: Bacterial > Pathogenic Escherichia coli infection.
· Human Diseases > Infectious diseases: Viral > Human papillomavirus infection.
· Human Diseases > Infectious diseases: Viral > HTLV-I infection.
· Human Diseases > Cancers: Overview > Pathways in cancer. (View pathway)
· Human Diseases > Cancers: Overview > Proteoglycans in cancer.
· Human Diseases > Cancers: Specific types > Colorectal cancer. (View pathway)
· Human Diseases > Cancers: Specific types > Endometrial cancer. (View pathway)
· Human Diseases > Cancers: Specific types > Prostate cancer. (View pathway)
· Human Diseases > Cancers: Specific types > Thyroid cancer. (View pathway)
· Human Diseases > Cancers: Specific types > Basal cell carcinoma. (View pathway)
· Human Diseases > Cancers: Specific types > Breast cancer. (View pathway)
· Human Diseases > Cancers: Specific types > Hepatocellular carcinoma. (View pathway)
· Human Diseases > Cancers: Specific types > Gastric cancer. (View pathway)
· Human Diseases > Cardiovascular diseases > Arrhythmogenic right ventricular cardiomyopathy (ARVC).
· Organismal Systems > Immune system > Leukocyte transendothelial migration. (View pathway)
· Organismal Systems > Endocrine system > Melanogenesis.
· Organismal Systems > Endocrine system > Thyroid hormone signaling pathway. (View pathway)
References

Application: WB Species: Mouse Sample:

Application: WB Species: human Sample:

Application: WB Species: Mouse Sample:

Application: WB Species: Mouse Sample:

Application: WB Species: Rat Sample:
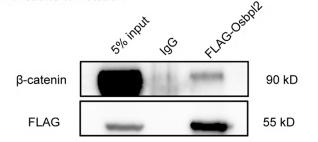
Application: WB Species: Human Sample: HEK293T cells
Restrictive clause
Affinity Biosciences tests all products strictly. Citations are provided as a resource for additional applications that have not been validated by Affinity Biosciences. Please choose the appropriate format for each application and consult Materials and Methods sections for additional details about the use of any product in these publications.
For Research Use Only.
Not for use in diagnostic or therapeutic procedures. Not for resale. Not for distribution without written consent. Affinity Biosciences will not be held responsible for patent infringement or other violations that may occur with the use of our products. Affinity Biosciences, Affinity Biosciences Logo and all other trademarks are the property of Affinity Biosciences LTD.